

n my earlier post on Gregory Clark’s work, The Son Becomes The Father, I laid bare the case for the known high heritability of human behavioral traits (including values and attitudes) and life outcomes. As well, equally important, I illustrated the complete absence of shared environment influences on these – that is, the effect common environmental forces that children growing up together share. This includes parents and upbringing, making it abundantly clear that parents don’t leave a lasting impact on who we grow up to be. These are towards what I’m calling the “75-0-25 or something” rule, which echos Satoshi Kanazawa’s “50-0-50 rule” summarizing behavioral genetic research, only more accurately so. This “75-0-25 or something” rule means the following:
For the variance in behavioral traits and life outcomes, each factor accounts for the following fractions:
- Heredity: 75% (though the variance is typically anywhere from 65% to almost 100%)
- Shared environment: 0%
- Unknown (classified as the “unique environment” or the “unshared environment”): 25%
About that third component, the unexplained variance, or the “unique environment,” we have thus far failed to reliably find anything that can account for it. It is deemed “environmental” in the sense that it is not heritable and presumably not genetic in origin (although that is not likely entirely accurate), but we have no idea what factors comprise it.
It’s important to note these values only represent the estimates within a generation. It doesn’t necessarily hold between generations (or between other contexts), where gross environmental changes can and do have large effects on the manifestation of human traits. See my post Why HBD. (Anyone who fails to absorb this gets an automatic F grade.)
However, the large heritability estimates and the failure to find any reliable environmental modulates of within-cohort differences casts doubt on most any environmental theory. Quite likely, as with the case with diet and lifestyle (see Tweet of the Week), most of the ones you’ve heard are bunk. It seems likely that the human genome draws up a design for a certain organism (that is more or less faithfully executed) that is designed to function to process and negotiate its environment in a certain set of ways, and precise behavior depends on the circumstances of the day and the specifics of the situation (see my post Environmental Hereditarianism). This may be as good of an explanation as any, but I am looking for evidence of reliable environmental modulation. One apparent environmental modulation is that some individuals with certain sensitive temperaments may be permanent modified by certain environmental exposure. Post-traumatic stress disorder (PTSD), for example, appears to be highly heritable. As well, it seems only certain individuals are susceptible, even after experiencing traumatic events. This illustrates that the susceptibility and extent of environmental modification of human traits is bounded by genetic prescription.
The high heritability of behavioral traits and the general lack of environmental effect on human behavioral traits (along with knowledge of the past and evolutionary theory) also leads one to understand that differences between human groups, be those groups be races, nationalities, ethnic groups, regional populations, even clans/families (as with Gregory Clark’s work), must also have some (rather large) genetic basis. “Culture” then is to be understood as the collective manifestation of those differences – the product of each individual’s innate propensity acting on each other’s and the circumstances of the day. Hence, genetic factors underlie these divisions:

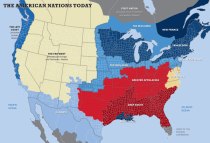

For the above reasons, this should be a given, but, we see how that is received in this day and age.
Even for those who accept heritable human differences, there is some discomfort at their full implications. My earlier post More Twitter Wisdom featured discussion of the unease many people have with this notion. Ironically, this itself is due to heritable factors (and yes, for those who can’t connect the dots, the ease that I have with this idea is in part the product of my genetic inheritance). Some people fight this notion tooth and nail, often eschewing logic in the process. Useful information can come out of these discussions, however, and this post will be an example. A recent troll commenter took that track, and here I will answer his claims.
Heritability, as established by behavioral genetics, comes from multiple lines of evidence, spanning completely different study methods. Most poignant today are Genome-wide Complex Trait Analyses (GCTA), which look at the correlation between resemblance in phenotype to directly measured resemblance in genotype. These have confirmed the (additive, anyway) heritable contribution to traits like IQ, BMI, and alcoholism, etc. However, even before these direct analyses came on the scene, the older methods firmly established the high heritability of behavioral and cognitive traits.
The bulk of behavioral genetic studies compare identical twins (monozygotic, MZ) raised together with fraternal (dizygotic, DZ) twins raised together. This is a particularly powerful method, because you control for a vast array of confounds, down to shared uterine environment. The difference in similarity between MZ twins and DZ twins estimates heritability. However, this method has weaknesses. It truly can’t rule out shared environmental influences as being behind the twin correlation. And further still, even to the extent that it produces a shared environment measure, it can’t separate the effect of assortative mating from this (which inflates the DZ twin correlation). You need confirmation from other methods. And such other methods exist.
The most “pure,” in terms of assessing heritability, are studies looking at twins raised apart (MZA and DZA, as opposed to MZT and DZT). Indeed MZA studies give you direct measurements of the heritability estimate. Additionally, there are adoption studies, which – when looking at adopted siblings – give you a direct measurement of the shared environment. Other pedigree studies (full siblings vs half-sibs, sibs vs. adoptees) buttress these. And finally, the extended twin design, which look at not just twins, but parents, spouses, cousins, etc. can allow you to separate a variety of influences.
One study gives a chart neatly summarizing the results of all of these for IQ:
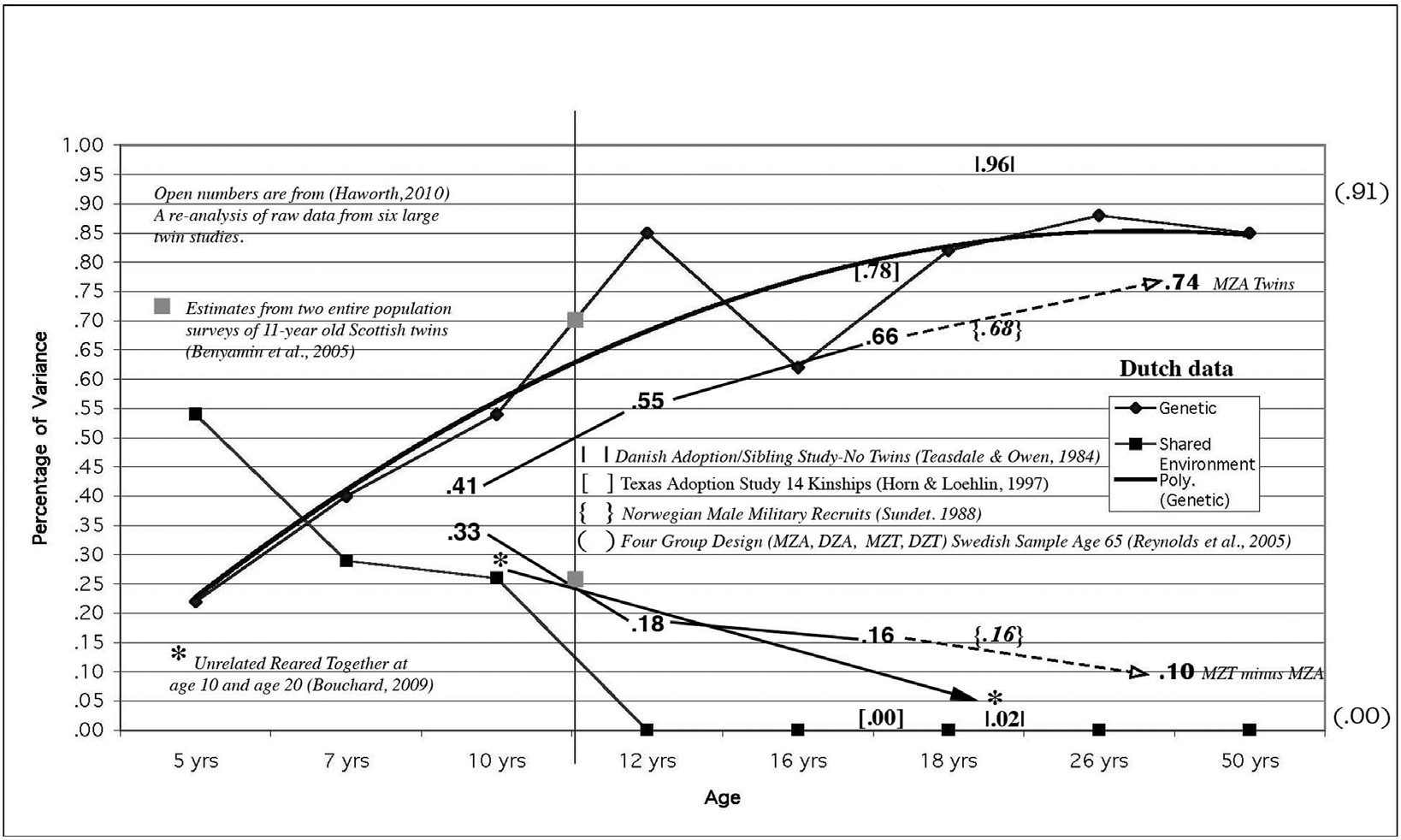 This is from The Wilson Effect: The Increase in Heritability of IQ With Age (2013). The heritability of IQ in adulthood from large samples of twins reliably returns in the 0.8 range. As we see here, a few studies, including a reasonably sized (n ~ 500) from Sweden that was a MZT-MZA-DZT-DZA study found a heritability of IQ of 0.91 at age 65. One study (a biological sibling-adoption study, small sample though) found a heritability pushing 100%!
This is from The Wilson Effect: The Increase in Heritability of IQ With Age (2013). The heritability of IQ in adulthood from large samples of twins reliably returns in the 0.8 range. As we see here, a few studies, including a reasonably sized (n ~ 500) from Sweden that was a MZT-MZA-DZT-DZA study found a heritability of IQ of 0.91 at age 65. One study (a biological sibling-adoption study, small sample though) found a heritability pushing 100%!
As seen here, adoption studies confirm the high heritability of adult IQ.
MZA studies, which are few and small, also confirm this. An apparent gap between the heritability found in MZA studies and the others is seen here. However, the combined sample size of MZA studies is only 187! The average heritability estimate from these of 0.74 is not horrendously off when one considers the small sample sizes.
The high early shared environment influence shows that in youth, environmental factors can make a difference. These influences diminish and disappear with time, dashing hopes of lasting parental influence. Some voices – including preeminent behavioral geneticist Robert Plomin himself – often try to claim that the increasing heritability of IQ and other behavioral traits can be boiled down to “gene-environment correlations” (rGE). The idea being that people seek out environments to suit their genetic proclivities (which they do), and the influence of that environment leads to the final trait. This is a nice rosy idea, because it appears to leave the door open to environmental manipulation, if we could intervene in the “proper” ways. However, it is fantasy. We clearly saw in my earlier post that the “gene-environment co-variance” was often negative! One’s environment seemed to be “making” one the opposite of what one would expect. Our experiences don’t shape our political attitudes like we think they do. So is the case with IQ.
Indeed, a meta-analysis of longitudinal twin and adoption studies attempted to test this idea. It sought to determine whether the increasing heritability of IQ could be explained by on-going environmental influence or genetic “amplification”; that is, the compounding of genetic effects over time. This is likely because the effect of each additional gene becomes more and more relevant as children grow up. Indeed, amplification is what they found:

From the first link:
Initially, the rGE explanation seems reasonable; when it comes to individuals differences, it doesn’t seem outlandish to suppose that naturally born intellectuals might increase their verbal IQ through bookish behavior. On the subpopulation level, likewise, it’s doesn’t seem implausible that a propensity for studiousness might lead to cognitive enhancement. Yet, there are a few bumps which preclude a simple rGE model. For one, it’s not merely this or that measure of intelligence that increases but, rather, the central factor and its numerous correlates [6]. In effect, rGE theorists are forced to maintain that g is created from the outside in. (See theoretical diagram below). Since g is structurally the same across individuals, cultures, sexes, and subpopulations, not only would the patterns of one’s environment have to construct g, the patterns of everyone’s environment would have to construct the same g.Additionally, IQ g has numerous endophenotypic correlates, such as the volume of white and grey matter, the mass of the prefrontal lobe, and total brain size [1, 11, 13] and the overlap between IQ (g) and many of these endophenotypes is entirely due to genetic influences [1, 13]. To explain this genetic covariation, rGE theorists must maintain that genetics sets the parameters for environmental selection, which leads to the development of different cognitive phenotypes, which, in turn, molds the endophenotypic differences, thus creating the three way correlation. Since the Phenotypic/endophenotypic correlations have been found to be a function of differential rates of change during the development process [9] (see figure 2), rGE theorists must maintain that this environmentally induced endophenotypic molding occurs primarily during the developmental process and starts early on. If we kept in mind what we said above, that the genes that lead to slight genetic IQ differences in infancy are the same genes that lead to large genetic based differences in adulthood, and note that the heritability of many dispositions also increases with age [4], we can readily identify the problem with this conception. Somehow, dispositional differences, which are under heavy environmental influence early on, must set the phenotypic/endophenotypic molding (environmental) parameters in a way that happens to correspond to the genetic driven phenotype that the individual will later express.
{kind=link}
Also, from the third link above:
All of this should now be a moot point since Davies et al. (2011) has established the high narrowheritability (i.e., breeding value) of IQ. (If the high heritability or increasing heritability found in conventional twin studies was a function of (passive/evocative/active) GE correlations, as was argued, Davies et al. (2011) would have found a much lower narrow heritability in their study.)
***End Edit***]
Another criticism of the heritability of IQ is that it is modulated by socioeconomic status (SES) – that IQ is less heritable in poor families. A large and extensive study co-authored by Robert Plomin reviewed the literature on the gene-SES interaction and conducted their own analysis of this phenomenon (with a sample of 8,716 twin pairs) and found no such effect. The heritability was constant across SES, but the shared environment vs. unique environment varied (this was done on children).
As well, for the record, though the overwhelming bulk of data for behavioral genetic studies comes from Northwestern European countries and their derivatives, behavioral genetic results have been gathered outside the Western world. The same results for the genetic and environmental influences on IQ has been found in Japan, for example (additive heritable influence, A ~ 0.8; shared environment, C = 0). This is in line with other behavioral genetic results generated there and in South Korea, as mentioned in my earlier post. Many behavioral genetic studies are underway outside the West, and we should have these results soon.
But on that point, another criticism leveled at behavioral genetic studies is that, in the United States at least, IQ is less heritable for racial minorities such as Blacks and Hispanics. However, this too is false. Over at Human Varieties, analyses by bloggers Chuck and Dalliard (also here) found no modulation of the heritability of IQ by race.
Proponents of the efficacy of nurture – especially parenting – often repeat a few erroneous arguments. Here I will address them. One of them is the idea that parenting, while ineffective for most, may make a difference for individuals with certain temperaments. For example, perhaps the low IQ/shiftless/delinquent/criminal or otherwise poorly dispositioned might benefit from more authoritative parenting, say. It’s a nice idea to think about, but it doesn’t happen. This is essentially “Stolen Generations” wisdom. As we’ve seen in my earlier post, a massive review of twin and adoption studies found no significant shared environment effect on criminality in adults (well, modeling found a shared environment contribution of 0.09, which can generally regard to be non-significant given the enormous measurement error expected). Even an effect that operated on some children but not others would contribute to the overall average shared environment, which was negligible.
Edit, 6/5/14: [I wanted to expand on the above mentioned review of criminality (by Rhee & Waldman, R&W), particularly the appearance of a small but nonzero (though non-significant) shared environment finding. As we saw, the age the subjects are assessed seems to make a difference. As well, as discussed in my analysis on adolescent psychopathology below, the particular measure used – such who is doing the ratings – affect the values found. For example, self-ratings or ratings by parents tend to attenuate the heritability estimate, and both appear to inflate the shared environment estimate, at least in youth. The Rhee & Waldman meta-analysis is no exception. Here are the ADCE (A, or a2 = additive genetic variance; D, or d2 = non-additive genetic variance; C, or c2 = shared environment; E, or e2 = remaining variance) components as computed based on information given by different raters:
| Rating method | a2 | d2 | c2 | e2 | Total no. of pairs in category |
| Self-report | 0.39 | – | 0.06 | 0.55 | 13,329 |
| Other report (usually parents) | 0.53 | – | 0.22 | 0.25 | 6,851 |
| Criminal records | 0.33 | 0.42 | – | 0.25 | 34,122 |
The total, or broad-sense heritability, H2 , is the sum of the additive (the narrow-sense heritability) and the non-additive genetic components. As we can see, when actual criminal records (a semi-objective metric) are used, as we’ve seen, the heritability shoots up to the usual range, at 0.75, and the shared environment estimate vanishes. The criminal record analysis also captures the largest number of subjects, bolstering its reliability. Parent reports, as seen below, inflate the shared environment measure. The self-report gives a negligible shared environment estimate, but reports a lower heritability estimate – which is not surprising, given that we can expect self-reported criminal behavior to be poorly reliable. It is unfortunate that R&W don’t separate out parents from peers and other non-relative raters in “other report.” Additionally, the adoption studies found a negligible shared environment impact of 0.05 between adoptive parents and adoptees. It is also too bad that R&W don’t cross tabulate the results by rating and age. But, as discussed below, adolescent shared environment effects maybe an artifact of unreliable raters anyway.
(For the record, the countries spanned by the studies in the meta-analysis include the U.S., Canada, the U.K., Australia, Denmark, and Sweden.)
The bottom line, it’s clear that when it comes to anti-social behavior, the 75-0-25 rule holds perfectly firm. Parents and parenting do nothing to create upstanding citizens, and heredity is considerably important. ***End Edit***]
Indeed, also supporting this is another massive meta-analysis of behavioral genetic influences on adolescent psychopathology (personality disorders). These captures various types of child misbehavior and dysfunction, including convenient diagnoses such as “oppositional-defiant disorder.” A look and the breakdown of their results is far more interesting than their main reported results. Typically, shared environment effects are seen in children (<18 years old). The main study reported this, but fortunately, they decomposed the type of measurements used. In addition to self-report and parental report, they also had teacher report, peer reports, and clinical diagnoses. The self and parental reports showed lower heritabilities (0.3-0.5) and significant (though small) shared environment components. However, when teacher or peer reports were used, they found much higher heritabilities, in the 0.65-0.8 range. As well, the shared environment impact vanished. Using clinical diagnosis also produced a zero shared environment impact. Considering the sheer size of this review, it’s clear that parental behavior dosen’t contribute to this malaise, even at these ages.
The problem of somewhat unreliable measurements (noise), especially coming from self-report, was illustrated in my earlier posts. Averaged peer ratings serve to adjust for this problem to an extent both by providing more proper social context by which to make accurate comparative ratings and by cancelling out fluke readings. Indeed, one behavioral genetic study, which attempted to investigate the idea of a “general factor of personality” (GFP), akin to g for cognitive ability, found that when using the combined scores of self and peer ratings, the heritabilities of the Big Five personality traits shot through the roof, with the additive heritable component being:
- Extraversion: 0.86
- Openness: 0.92
- Neuroticism: 0.59
- Agreeableness: 0.85
- Conscientiousness: 0.81
This demonstrates that more accurate measurements consistently push up the heritability estimate (even pushing them towards 100%), giving us the basis of the 75-0-25 or something rule.
As for the sixth dimension of personality, “honesty-humility”, the H component of the six factor HEXACO, evidence of its high heritability is also established, as we saw previously. Indeed, a recent post by Peter Frost (Evo and Proud: Compliance with moral norms: a partly heritable trait?) discussed a twin study from Sweden that looked at various forms of dishonesty, such as fraudulently claiming sick benefits or evading taxes. And sure enough, these particular behaviors showed considerable heritability. There is a desperate need for cross cultural behavioral genetic analyses. Many dimensions of personality systems like the HEXACO (as imperfect as they are) are likely to systematically vary from culture to culture.
The usefulness of behavioral genetics – indeed, the single most powerful and solid area of all social science – is highly evident. But behavioral genetic methods can be used to address several long-standing questions. Here we see it’s clear that parents don’t leave much of an impact on our behavioral traits. But what about people who aren’t parents? Here I will look at two sets of important people, spouses and peers.
It is no secret that spouses correlate on behavioral traits. This, assortative mating, is a powerful force, as we’ve seen previously. There are two aspects where spouses are highly correlated – the things you don’t talk about in a bar: politics and religion. Some have assumed that a good bit of this is because spouses grow more similar with time. But is this the case?
This is where the “extended twin” design comes in handy. One large study (N > 20,000) in particular looked at precisely that. By including twins, their spouses, and parents, etc, they were able to directly measure assortative mating. What did they find? Spouses were correlated for several traits. But the traits they were most correlated in were political orientation and religiosity. Social “homogamy” (having the same background as your spouse) couldn’t explain this, as the correlation between MZ twins and their co-twin’s spouse were consistently higher than that of DZ twins, and so on. As well, spouses weren’t influencing each other, as the correlation between spouses was not affected by length of the marriage (even when only couples married <2 years were examined).
The study was also able to lay to rest another persistent myth. We’ve heard that we choose spouses like our opposite sex parent (like our mothers for men and like our fathers for women). Anyone who’s remotely genetically informed should be able to see that this could just be due to choosing mates like ourselves. And so is the case. As the authors put it:
there was no evidence for the sexual imprinting hypothesis. Twins’ partners were not significantly more similar in any trait to the twin’s opposite-sex parent than to the twin’s same-sex parent or a DZ co-twin of either sex, nor was there even a trend in this direction
These results were also consistent with the Peter Hatemi et al extended twin study on political attitudesfeatured previously.
The similarity between spouses has nothing to do with mutual influence, but assortment. At least this bit is common sense. I suspect few long married individuals will believe that they changed their spouse.
On that note, a key theory put forward by the woman who first elucidated the non effect of parents, Judith Rich Harris, was that the unique environment “influence” might be boiled down to peer influence. Staffan did a nice recap of Harris’s theory (see The Nurture Enigma – How Does the Environment Influence Human Nature? | Staffan’s Personality Blog). We all have heard of peer pressure. And indeed, peers seem to be an important force when it comes to language and behaviors like smoking initiation. But do peers really have this great influence, as Harris posits? Well, as I posted over at the Lion of the Blogosphere:
Most research into peer effects is confounded by the same thing that standard parenting studies are: inability to control for the effect of heredity.
And:
A behavioral genetic study (on the Add Health data) that looked specifically at GPA and found that 72% of the similarity between U.S. high school students and their peers could be explained by genetic factors. In other words, school performance and the apparent peer “influence” is really just kids choosing to associate with kids of similar intelligence and motivation:Peers seem like a fine avenue to get excited about, because it seemed like a vehicle through which parents could assert some influence. But, when you really consider it, peers can’t really be all that important in the long run, because if there were systematic effects of peers on adult outcomes, it’d turn up in the shared environment, which it doesn’t. One could posit that the effect of peers is completely random, but if that were true (aside from the major violation of Occam’s Razor that presents), why worry about it?
The “75-0-25 or something” rule is robust and reliable. This instructs that should we find some major deviation from this, it can be taken to be a sign something is seriously amiss. We saw that with male homosexuality (see Greg Cochran’s “Gay Germ” Hypothesis – An Exercise in the Power of Germs). Now I will discuss two curious exceptions to this pattern.
One rather astonishing example was the heritability of social trust. A behavioral genetic study out of the Netherlands found that the heritability of trust in others, as measured by:
The trust-in-others and trust-in-self scales were designed to include three items that were central in existing scales … thereby capturing items with positive valence (“I completely trust most other people”) and negative valence (“When push comes to shove, I do not trust most other people”), both of which explicitly used the word “trust”, and an item that captured the broad behavioral implication of the trust: the intention to accept vulnerability, as explicated in one of the most widely-accepted definitions of trust … (“I dare to put my fate in the hands of most other people”)
…found no significant heritable influence on these. The extent that people trusted, at least as captured by these measures, was virtually entirely a function of the unique environment.
This was a puzzling result. The clear pattern of the high heritability of all behavioral traits was established, as I’ve discussed. So how could a propensity to trust not also be influenced by genetic factors? One explanation touted around was that trust is contingent on experience; if we found people trustworthy, we would trust. If we didn’t, we would not. While that might sound convincing, the trouble is that the same could be said for many other behavioral traits. Is general trust less socially contingent than say bigoted feelings against some groups, like homophobia (which is at least 54% heritable)? That seems rather unreasonable.
One key question: how do they assess “trust”? Just how good was their measurement? Measurements in social science need to meet three basic criteria: they need to be reliable (that is multiple testing instances of the same individual should give roughly the same results), they need to be “valid” (that is, be predictive of some real-world outcome), and they should be heritable. This trust measure clearly fails on the third criterion. However, the study authors claim the test-retest correlation was good, so it is reliable. But what about the second? Does this trust measure actually predict anything?
To find out, I looked at a study that sought to answer that very question. This study, done in Germany, looked in detail at the reliability and the validity of their measurement of trust, a measurement very similar to the Dutch study. The noted a key point, one HBD Chick will appreciate. That is, trust is multi-faceted. There is trust in institutions, which is distinct from trust in known others, which is distinct from trust in strangers (I’d imagine HBD Chick would break it down one more, and separate “known others” into family and non-family). But more importantly, to question of validity, they assessed this by the correlation between trust in strangers and trusting behavior in the “dictator game.” They found a correlation, but only with trust in strangers.
But their correlation was very small (Spearman’s  = 0.17) – and this is with a game which itself has questionable relation to trust behavior in the real world. I suspect that their instrument is not predictive of any trusting behavior in the real world. It’s worth mentioning another (fairly small) study of the heritability of trust from Australia found a non-insignificant heritability, though a smallish one (0.14-0.31).
= 0.17) – and this is with a game which itself has questionable relation to trust behavior in the real world. I suspect that their instrument is not predictive of any trusting behavior in the real world. It’s worth mentioning another (fairly small) study of the heritability of trust from Australia found a non-insignificant heritability, though a smallish one (0.14-0.31).
= 0.17) – and this is with a game which itself has questionable relation to trust behavior in the real world. I suspect that their instrument is not predictive of any trusting behavior in the real world. It’s worth mentioning another (fairly small) study of the heritability of trust from Australia found a non-insignificant heritability, though a smallish one (0.14-0.31).
The situation with trust is unclear. But this brings me to another example of a feature for which the heritability estimate appears to be trivial. That is the female G-spot. A study on about 1,800 female twins from Britain found that the heritability of the reported presence of a G-spot wasn’t significant. The result was virtually entirely unshared environment. Debate has raged on as to whether or not the female G spot exists at all, but that is to be expected, since research into human sexual behavior is among the most difficult to conduct properly. But, the result from this study indicating that the G spot isn’t heritable is puzzling. If the G spot was a real anatomical feature, and one that wasn’t universal, then one would expect a rather significant heritable impact. The finding that it’s not heritable points to one of two conclusions. One, perhaps the G-spot is in fact universal, but only some women have “discovered” it. That seems rather implausible, given the rather significant variation in heritable morphological features of sex organs in women. The second possibility is that the G-spot in fact doesn’t exist at all, and women who claim to have one are mistaken. That seems more likely, but I wouldn’t want to completely dismiss the claims of women who state they have such a feature. The mystery remains.
The findings of behavioral genetics, particularly the highly significant impact of heredity and the absence of shared environment effects, in addition to the complete failure to find reliable environmental sources that contribute to the “unique environment” component of the variance, calls into question virtually every pet environmental theory that has been put forward. It guides one to be suspicious of most “environmental” explanations of behavior. Now, let me be clear, I am not saying that these environmental influences don’t exist. I am not saying that if they do exist, we won’t be able to ever find them. I am also not saying that development doesn’t require a complex interplay between genes and environment. Try going without food, water, air, or speaking to another person if you don’t believe me. I am also not saying that the secular changes in human traits that are brought about by gross environmental changes don’t happen. The increase in average height over the past century disproves that. But what I am saying is that you should be doubtful of most pet theories of how the environment influences us, especially those that promise we can control, or sometimes even predict it. For as we see, that’s far from an easy task.
(Source: jaymans.wordpress.com)

No hay comentarios.:
Publicar un comentario